
药用植物可以预防、治疗疾病,具有极高医疗和经济价值,中国劳动人民几千年来在长期和疾病作斗争的过程中,对药用植物的使用积累了丰富的经验[1-2]。药用植物的药效活性成分主要为其次生代谢产物,研究药用植物次生代谢物产生过程、代谢途径、调控方式等是药用植物研究的重要组成部分[3]。青藏高原是一个天然的药用植物资源库,据统计约有1 200种药用植物,分布于青藏高原的药用植物生命力极强,具有耐寒耐旱、有效成分积累高、生物活性强等特点[4]。红景天Rhodiola rosea Linn.、珠芽蓼Polygonum viviparumLinn.和瑞香狼毒Stellera chamaejasme Linn.是青藏高原常见的3种药用植物,在中药和民族药中具有重要的药用和经济价值。
红景天R. rosea L.为景天科(Crassulaceae)红景天属RhodiolaL.多年生草本药用植物,主要生长在青藏高原海拔3 000 m以上的高寒地区[5]。红景天在我国具有悠久的药用历史,其主要活性成分有红景天苷(salidroside)、酪醇(tyrosol)、红景天素(rhodosin)、肉桂醇苷(rosavin)和草质素苷(rhodionin)等,具有抗缺氧、抗辐射、保护心脑血管、抗肿瘤与增强免疫力等多种功效[6]。珠芽蓼P.viviparum L.为蓼科(Polygonaceae)蓼属polygonumL.多年生草本药用植物,主要分布于青藏高原高山草甸,全株具有广泛的药用价值和开发前景。珠芽蓼含有挥发油、黄酮、多糖、酚酸类成分,具有清热解毒、消肿止血、抗肿瘤等功效,是常用的中药材[7]。瑞香狼毒S. chamaejasmeL.为瑞香科(Thymelaeaceae)狼毒属StelleraL.多年生草本药用植物,广泛分布于青藏高原土壤退化的草地[8]。瑞香狼毒的根作为传统中药,主要活性成分有香豆素类、黄酮类、二萜类、木脂素类化合物,具有抗肿瘤、抗菌、抗病毒、杀虫、抗炎、调节免疫等生物功能[9]。
植物根系不断分泌各种代谢产物,这些代谢产物进入土壤,使得根际土壤的营养物质和微生物比较丰富,并且其理化性质和生物学特性与非根际土壤存在非常大的差别[10]。植物根系分泌物的主要成分为各种各样的含碳代谢物、各种酶、离子和水等,在调控根际土壤环境的植物生长、营养平衡和生物化学循环等方面发挥着重要作用,而且根系分泌物中不同的化合物、酶或某些特异代谢产物对植物和根际微环境会产生特定影响[11]。因此,对植物根际土壤代谢产物的研究,有助于揭示植物种群与土壤环境之间的信息传输和物质互换,为研究植物对自然环境改变的响应和调控养分转换等方面奠定基础。
非靶向代谢组学可以全面、系统地对机体、组织器官等中所有相对分子质量在1 000以内的小分子代谢同时进行定性和定量分析,能尽可能完整客观地表征样品中代谢物种类,关注生物学样本之间的相似性、聚类情况及差异变化[12]。超高效液相色谱-质谱(UPLC-MS)联用技术在检测和量化植物根际土壤代谢产物方面被广泛利用,该技术分离性好,灵敏度高,可以快速准确地分析样本内所含有的代谢物,结合多元统计方法筛选差异代谢物,并预测代谢物可能的代谢途径和代谢机制[13]。近年来非靶向代谢组学技术已广泛地应用于天然产物质量控制与评价、作物品质改良、资源鉴定、新生物标记物开发等[14-16]。然而,对于青藏高原高寒草地药用植物根际土壤代谢组学方面的研究相对匮乏。
甘肃省武威市天祝藏族自治县地处祁连山东端,属青藏高原东北边缘,天然草地资源丰富,主要分布在抓喜秀龙、松山、毛藏、西大滩等地。天祝县抓喜秀龙乡天然草原土壤有机质含量高,祁连山脉的终年积雪提供了丰富的降水资源,在抓喜秀龙草原分布着多种耐寒药用植物[17]。研究表明,大多数野生药用植物受地理环境和气候因素等的影响,其药效品质高于栽培药用植物[18]。红景天、珠芽蓼和瑞香狼毒是重要的藏族药用植物[17],为高寒地区藏族人民的医药产业发展发挥了重要的作用,也是民族植物资源的重要组成部分。红景天、珠芽蓼和瑞香狼毒在天祝县天然草地广泛分布,且互为伴生植物,这些药用植物资源在当地藏族文化和生活中有很高的地位。然而,目前当地药用植物利用率低,野生药用植物资源没有得到合理有效的开发利用。因此,对当地广泛分布的野生药用植物资源进行深入研究,既对藏族药用植物的保护与开发具有积极推动作用,还能维护当地民族植物资源的有效利用和可持续发展[17]。
为了比较天祝县抓喜秀龙乡高寒草原广泛分布的红景天、珠芽蓼、瑞香狼毒3种药用植物根际土壤代谢产物,揭示其适应高寒草原生态环境的物质基础,本研究以这3种药用植物的根际土壤为实验材料,采用高分离度液相色谱-四极杆飞行时间质谱联用技术(UPLC-QTOF-MS)的非靶向代谢组学方法,分析研究3种药用植物根际土壤代谢物的组成和特性;基于主成分分析(principal component analysis,PCA)、聚类分析、热图分析、通路分析等多元统计分析方法,筛选出3种药用植物的差异代谢物,并分析其代谢通路,有助于揭示青藏高原高寒草地不同药用植物的根际过程,为药用植物适应高寒草地环境及根际养分转化等提供理论依据。
1 材料与仪器
1.1材料
采样地点设置在甘肃省武威市天祝藏族自治县抓喜秀龙乡(N37°09′39″,E102°48′22″),海拔高度为2 874 m,采集时间为2021年7月15~18日。3种药用植物的标本由兰州大学生命科学学院的潘健斌讲师分别鉴定为瑞香狼毒S. chamaejasme Linn.、珠芽蓼P. viviparumLinn.和红景天R. rosea Linn.,保存在西北民族大学化工学院。选择瑞香狼毒、红景天和珠芽蓼生长盖度较高的样地,设置3个10 m×10 m的样方,每个样方间隔50 m以上。在每个样方内,分别采集8株大小、长势一致的瑞香狼毒、红景天以及珠芽蓼植株,抖落根部的土壤,将粘附在根部2 mm内的土壤,用干净毛刷将所有土壤收集到采样袋内,即为根际土壤。将每个样方每种植物得到的8份根际土壤分别进行均匀混样,即得到瑞香狼毒、红景天和珠芽蓼的根际土壤各3份。分别取5 g左右的土壤放入10 mL离心管,并用封口膜封口,液氮速冻10~15 min,−80 ℃保存备用。
1.2仪器
高分辨质谱仪(赛默飞世尔科技公司);Nexera 型高效液相色谱仪(日本岛津公司);TYXH-I型漩涡振荡器(上海汗诺仪器有限公司);SB-5200DT型超声波清洗机(宁波新芝生物科技有限公司);TGL-16MS型台式高速冷冻离心机(上海卢湘仪离心机仪器有限公司);JXFSTPRP-24/32型全自动样品快速研磨仪(上海净信实业发展有限公司)
2 方法
2.1 代谢组样品前处理
实验前提取试剂甲醇、甲酸、水、乙腈以及L-2-氯苯丙氨酸均在−20 ℃下预冷,化学药品和溶剂均为色谱级。准确称取瑞香狼毒、红景天以及珠芽蓼根际土壤样品各0.50 g,加入内标(L-2-氯苯丙氨酸,0.3 mg/mL,甲醇配制)10 μL和1 mL甲醇水溶液(1∶1);依次加入2颗小钢珠,在−20 ℃放置2 min预冷,加入研磨机研磨(60 Hz,2 min);将匀浆后的样品转移至15 mL离心管中,用1 mL甲醇水溶液(1∶1)转移管壁残留,重复上述操作1次;离心10 min(4 ℃、7 700 r/min),取上清2.5 mL于5 mL离心管中,冻干;用400 μL甲醇水溶液(1∶4)复溶,涡旋震荡60 s,超声30 s;离心10 min(4 ℃、12 000 r/min),用注射器吸取150 μL的上清液,0.22 μm的有机相针孔过滤器滤过后,转移到LC进样小瓶,−80 ℃下保存,直到进行LC-MS分析。质控样本(quality control,QC)由所有样本的提取液等体积37.5 μL混合制备而成,QC的体积与样本相同;重复3次,在仪器检测的过程中,每6个检测分析样本中插入1个QC样本,以考察整个分析过程的重复性。
2.2 色谱条件
采用Nexera UPLC超高效液相串联QE高分辨质谱仪组成的液质联用系统对样品进行分离。分离条件:色谱柱:Waters,ACQUITY UPLC HSS T3(100 mm×2.1 mm,1.8 μm);流动相为0.1%甲酸水(A)-乙腈(B);梯度洗脱程序:0~2 min,95% A;2~4 min,95%~70% A;4~8 min,70%~50% A;8~10 min,50%~20% A;10~14 min,20%~0 A;14~16 min,0~95% A;柱温25 ℃;体积流量0.35 mL/min;进样量2 µL,整个分析过程中样品置于4 ℃自动进样器中。
2.3 质谱条件
样品质谱信号采集分别采用正负离子扫描模式,质谱扫描的选择范围为125~1 000 m/z,全扫描分辨率为70 000,高分辨高能量碰撞解离质谱(HCD MS/MS)扫描分辨率为17 500,毛细管温度为320 V。ESI 源设置为:检测模式ESI+,喷雾电压3.5 kV,鞘气体流量40 arb,辅助气体流量10 arb;检测模式ESI−,喷雾电压−3 kV,鞘气体流量35 arb,辅助气体流量8 arb。
2.4 数据处理与分析
2.4.1代谢物鉴定将得到的原始数据转换成mzXML格式,利用Waters公司的Progenesis QI v2.3软件,对原始数据进行预处理,将采集得到的谱图进行过滤噪音、解卷积、峰强度校正、峰定性和定量等分析,删除组内缺失值>50%的离子峰,最后整理成二维数据矩阵形式,包含保留时间(retentiontime,tR)、代谢物名称(metabolites)、质荷比(m/z)、化合物分子式(formula)等信息,再基于The Human Metabolome Database(HMDB)、Lipidmaps(v2.3)和METLIN数据库以及上海欧易生物医学科技有限公司自建库,质量误差设置为小于1×10−6,并结合二级质谱匹配得分,对代谢物进行鉴定。
2.4.2数据处理利用软件SIMCA-P 14.1进行多元统计分析,包括PCA、偏最小二乘法分析(PLS-DA)、正交偏最小二乘法分析(orthogonal partial least squares-discriminant analysis,OPLS-DA)。同时进行Student’s t-test和变异倍数分析,使用R 软件绘制火山图的单维统计分析方法。差异化合物筛选条件:P<0.05、VIP>1。利用R软件对VIP值前50差异代谢物的表达量进行层次聚类和绘制火山图对P值、VIP和Fold change值进行可视化,利于筛选差异代谢物。相关性分析使用Pearson相关系数衡量两个代谢物之间的线性相关程度。利用差异代谢物的KEGG ID进行通路富集分析,通过超几何检验,分析在显著性差异表达代谢物中显著富集的代谢通路(P≤0.05)。
3 结果与分析
3.1 质量控制和PCA
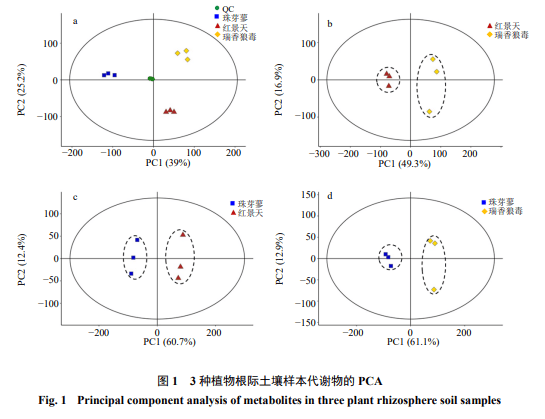
QC样本用于测定进样前仪器状态及平衡色谱-质谱系统,并用于评价整个实验过程中系统稳定性。采用XCMS软件提取代谢物离子峰,将提取得到的所有实验样本和QC样本的峰,标准化后经Pareto-scaling处理后进行PCA分析(图1-a)。经7-fold cross-validation(7次循环交叉验证)得到的总样品PCA模型参数,观察样本之间的分布趋势。QC样本紧密聚集在一起,表明此实验过程中仪器检测稳定性较好,本实验的重复性好。因此,本实验的质谱分析系统稳定性较好,得到的实验数据稳定可靠,可为接下来的实验结果提供有效参数。3种植物的所有根际土壤样本能够鉴定出的代谢物种类总有5 882种,其中正离子模式下有4 308种,负离子模式下有1 574种。对3种植物的根际土壤样本两两组合,分别进行PCA分析(图1-b~d)。从图1可以看出,无论正、负离子模式下,2组样本均处于各自的95%的置信区间内,3种植物的根际土壤均能显著区分。
3.2 根际土壤样本的OPLS-DA
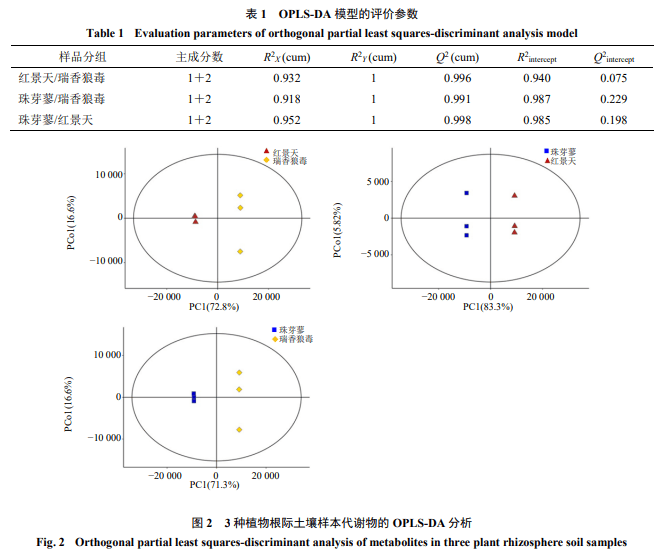
建立瑞香狼毒、珠芽蓼、红景天3种植物根际土壤样本分组的OPLS-DA模型(表1、图2)。由表1的模型评价参数可知,R2Y等于1,R2X、Q2均接近1,说明建立的模型稳定可靠。图2显示,3组样本中,瑞香狼毒和珠芽蓼、珠芽蓼和红景天、红景天和瑞香狼毒中,样本间均没有重叠,说明3种植物的组间差异较为明显。
3.3根际土壤样本差异代谢物的火山图分析
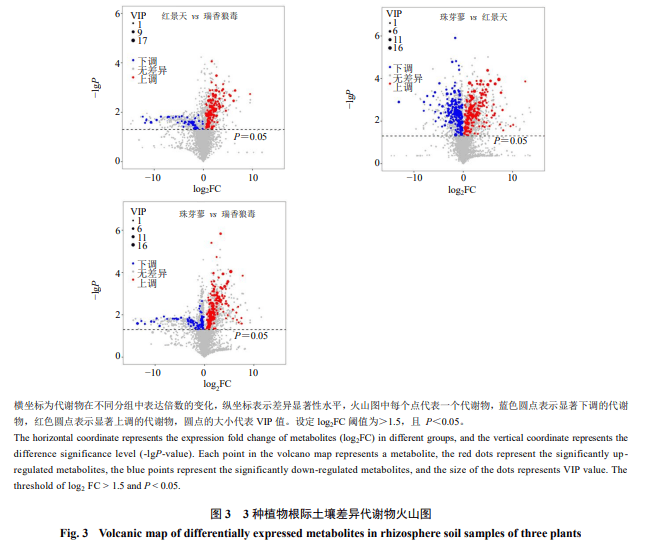
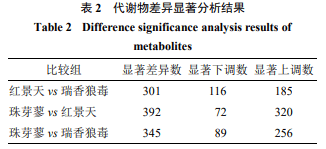
3种植物根际土壤样本两两组合差异代谢物火山图见图3,可知不论是那2种植物根际土壤样本,上调的差异代谢物比下调的代谢物都要高的多,尤其是珠芽蓼和红景天的根际土壤中显著上调的代谢物比下调代谢物高248种;瑞香狼毒和珠芽蓼根际土壤上调的差异代谢物比下调的代谢物高167种,瑞香狼毒和红景天根际土壤上调的差异代谢物比下调的代谢物高69种。从表2可以看出,瑞香狼毒和红景天根际土壤差异代谢物种类较少,上调的差异代谢物为185种,下调的代谢物为116种;珠芽蓼和红景天根际土壤差异代谢物种类较多,上调的差异代谢物为320种,下调的代谢物为72种;珠芽蓼和瑞香狼毒根际土壤差异代谢物种类也相对较多,上调的差异代谢物为256种,下调的代谢物为89种。
3.4根际土壤样本差异代谢物聚类热图分析
为了更直观的展示根际土壤样本之间的关系及代谢物在不同样本之间的表达差异,对各比较组显著差异代谢物VIP值前50(top 50)差异代谢物进行统计,并对前50差异代谢物的表达量进行层次聚类,聚类图见图4。图中红色为高表达代谢物,蓝色为低表达代谢物,颜色从蓝到红表示代谢物的表达丰度从低到高,即越红表示差异代谢物的表达丰度越高。3种植物根际土壤样本在两两比对时,代谢物均各自聚集在同一簇中,每个样品组代谢物在丰度上均差异显著,说明选择的差异代谢物可作为标志物将3组样品两两区分开来,其中有些代谢物聚集在一块,可能它们具有相似的功能或参与同一条代谢通路(图4)。
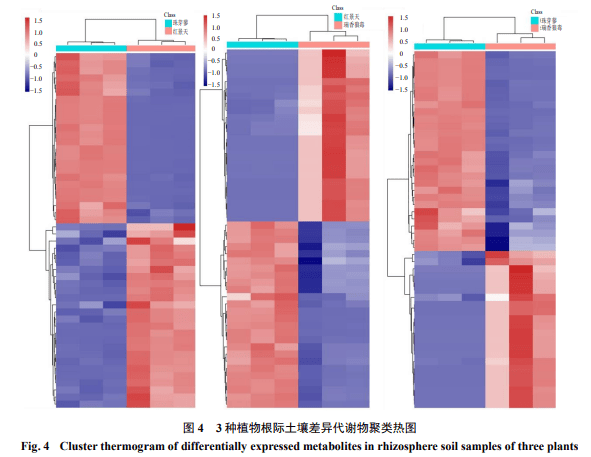
虽然50种显著差异代谢物在2种不同植物根际土壤上调或下调差异程度不同,但趋势同步。珠芽蓼和红景天根际土壤样本比较组中,50种显著差异代谢物可聚分为2大类:一类在珠芽蓼根际土壤高表达的,有24种;另一类为在红景天根际土壤高表达的,有26种(图4-a)。瑞香狼毒和红景天根际土壤样本比较组中,50种显著差异代谢物也可聚分为2大类:一类在红景天根际土壤高表达的,有24种;另一类为在瑞香狼毒根际土壤高表达的,有26种(图4-b)。同样地,瑞香狼毒和珠芽蓼根际土壤样本比较组中,50种显著差异代谢物聚分为2大类:一类在珠芽蓼根际土壤高表达的,有28种;另一类为在瑞香狼毒根际土壤高表达的,有22种(图4-c)。
3.5 根际土壤样本标志代谢物筛选
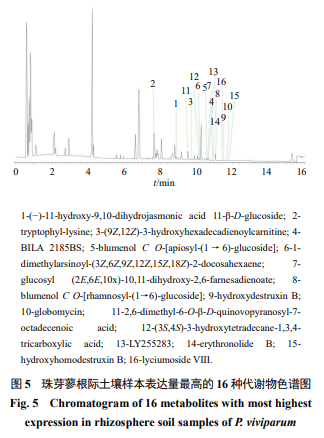
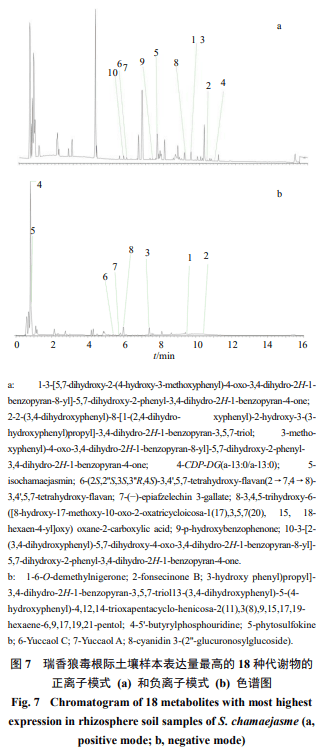
对瑞香狼毒、红景天和珠芽蓼根际土壤样本各比较组显著差异代谢物VIP值前50且P<0.05的差异代谢物的丰度聚类图进行比较,分别确定3个根际土壤样本比较组表达量最高的差异代谢物(表3)。珠芽蓼根际土壤样本中检测到表达量高的16种差异性代谢物,均在正离子模式下,而且所有差异性代谢物均上调,这些代谢物多属于烃、脂肪酸类、氨基酸类和糖苷类化合物(表3、图5)。红景天根际土壤样本中检测到表达量高的16种差异性代谢物(图6),正离子模式下有11种,负离子模式下有5种,所有差异性代谢物均上调,这些代谢物它们多属于碳水化合物及其结合物、氨基酸、肽类、糖苷类(图6、表4)。瑞香狼毒根际土壤样本中检测到表达量高的18种差异性代谢物(表5、图7),正离子模式下有10种(图7-a),负离子模式下有8种(图7-b),所有差异性代谢物均下调,这些代谢物多为双黄酮类、植物磺肽素、呋喃类、狼毒素类以及苯甲酮类化合物。




3.6根际土壤样本代谢通路富集分析
通过对3种植物根际土壤样本差异代谢物进行通路富集分析,可对各比较组样本中代谢途径的变化进行了解。基于KEGG数据库,利用差异代谢物的KEGG ID获得代谢通路富集结果,通过超几何检验,在显著性差异表达代谢物中显著富集的pathway条目,以此对差异代谢物进行代谢通路富集分析。亚油酸代谢(linoleic acid metabolism),甘氨酸、丝氨酸和苏氨酸的代谢,D-谷氨酰胺和D-谷氨酸代谢(D-glutamine and D-glutamate metabolism),嘌呤代谢(purine metabolism)4条代谢通路是3个比较组中极显著富集的代谢通路(P<0.01)。由代谢通路气泡图可知(图8),各比较组中ATP结合盒转运体(ABC transporters)、D-谷氨酰胺和D-谷氨酸代谢和嘌呤代谢3种代谢通路在图中点均较高,说明其代谢物相对较多,显著性较高,是三者重要的代谢通路。
4 讨论
次生代谢物是药用植物发挥药效价值的活性物质基础,药用植物通过生物合成,产生千变万化的分子结构以及多种多样的活性作用的次生代谢产物,是药物分子的重要实体来源。近年来代谢组学广泛应用于药用植物研究,为药用植物开发利用和资源保护发挥了重要作用[19]。植物根际土壤作为植物根系与外界土壤物质交换和能量流动的重要媒介,所占的体积相对较小,但植物根系不断地分泌各种代谢产物,以及根表组织持续凋亡脱落进入根际土壤,使得根际土壤的营养物质、微生物比较丰富,且其理化性质和生物学特性也有别于非根际土壤[20]。因此研究药用植物根际土壤代谢物的组成、差异代谢物分析、标志代谢物筛选以及代谢通路富集分析对深入研究药用植物次生代谢产物合成路径,调控因子以及代谢网络具有重要作用。
红景天、珠芽蓼和瑞香狼毒是青藏高原常见药用植物,也是民族药的重要组成部分。本研究发现,虽然3种药用植物在青藏高原具有共同的自然生活环境,但其根际土壤代谢物组成存在显著差异,PCA与OPLS-DA分析均表明,3种药用植物根际土壤样本间均没有重叠,组间差异较为明显。这可能与3种植物属于不同的科,亲缘关系较远有关。在对3种药用植物根际土壤样本显著差异代谢物分析中发现,珠芽蓼与红景天和瑞香狼毒的显著差异代谢物均比较多,而红景天和瑞香狼毒之间显著差异代谢物相对较少,这个结果在一定程度反映了珠芽蓼次生代谢产物的代谢合成途径可能更多变。
本研究发现,3种药用植物根际土壤样本的标志代谢物与其主要活性成分密切相关红景天根际土壤样本中检测到表达量最高的标志性代谢物多属于碳水化合物及其结合物、氨基酸、肽类、糖苷类等,碳水化合物及其结合物、氨基酸和糖苷类物质等是合成红景天苷、红景天素(主要为黄酮类物质)及红景天多糖的前体[6]。瑞香狼毒根际土壤样本中检测到表达量高的标志性代谢物多为双黄酮类、植物磺肽素、呋喃类、狼毒素类以及苯甲酮类化合物,而瑞香狼毒根中的多种黄酮类、香豆素类及二萜类等活性成分都具有较强的抗肿瘤、抗病毒作用[9],瑞香狼毒根部合成的活性化合物以根系分泌物的方式进入根际土壤,有利于瑞香狼毒种群的种间竞争和环境适应能力。珠芽蓼根际土壤样本中检测到的标志性代谢物多属于烃、脂肪酸类、氨基酸类和糖苷类化合物,据报道,珠芽蓼根茎富含挥发油、黄酮苷、香豆精、有机酸和多糖等活性成分[7],这些活性成分的生物合成前体和中间体中包括了其分泌到根际土壤中的标志性代谢物。由此可见,3种药用植物的某些活性次生代谢产物在体内合成之后,可通过根系分泌的方式进入根际土壤环境,影响药用植物的根际微域环境[21]。
在3种药用植物根际土壤样本的3个比较组中,ATP结合盒转运体(ATP transporters)、D-谷氨酰胺和D-谷氨酸代谢(D-glutamine and D-glutamate metabolism)、嘌呤代谢(purine metabolism)3种代谢通路的显著性最高,其中D-谷氨酰胺和D-谷氨酸代谢的富集程度最大。在3个比较组中,氧戊二酸(oxoglutaric acid)、聚-g-D-谷氨酸(poly-g-D-glutamate)存在于D-谷氨酰胺和D-谷氨酸代谢通路且显著上调,即植物体中D-谷氨酰胺在谷氨酰胺酶以及天冬酰胺酶催化下,合成L-谷氨酸,在谷氨酸脱氢酶催化下形成氧戊二酸,或直接转化为聚-g-D-谷氨酸,增加植物对氮素的利用率[22-23]。ATP结合盒转运体是植物体内重要的转运体家族,参与着多种代谢产物的分布,包括无机离子、单糖、聚糖、胆固醇、磷脂、氨基酸、肽、蛋白质、毒素、药物、抗生素和异源物质等[24-25]。嘌呤代谢(purine metabolism)是核酸碱基腺嘌呤及鸟嘌呤等的嘌呤衍生物的活体合成及分解的主要代谢途径[26-27]。这些代谢途径可能是不同药用植物在青藏高原高寒环境下,次生代谢产物生物合成、适应高寒草地生态环境、种间生态竞争及根际养分和能量循环的重要基础。
利益冲突所有作者均声明不存在利益冲突
参考文献(略)
来 源:程锦春,戴 璐,王丽红,黄先成.基于非靶向代谢组学分析3种青藏高原药用植物根际土壤代谢产物特征 [J]. 中草药, 2024, 55(8): 2734-2745.
- 本文固定链接: https://maimengkong.com/zixun/1726.html
- 转载请注明: : 萌小白 2024年5月2日 于 卖萌控的博客 发表
- 百度已收录
